 -
Volume 152,
Issue 7,
2006
-
Volume 152,
Issue 7,
2006

Volume 152, Issue 7, 2006
- Pathogens And Pathogenicity
-
-
-
Proteus mirabilis isolates of different origins do not show correlation with virulence attributes and can colonize the urinary tract of mice
More LessProteus mirabilis has been described as an aetiological agent in a wide range of infections, playing an important role in urinary tract infections (UTIs). In this study, a collection of P. mirabilis isolates obtained from clinical and non-clinical sources was analysed in order to determine a possible correlation between origin, virulence factors and in vivo infectivity. Isolates were characterized in vitro, assessing several virulence properties that had been previously associated with P. mirabilis uropathogenicity. Swarming motility, urease production, growth in urine, outer-membrane protein patterns, ability to grow in the presence of different iron sources, haemolysin and haemagglutinin production, and the presence and expression of diverse fimbrial genes, were analysed. In order to evaluate the infectivity of the different isolates, the experimental ascending UTI model in mice was used. Additionally, the Dienes test and the enterobacterial repetitive intergenic consensus (ERIC)-PCR assay were performed to assess the genetic diversity of the isolates. The results of the present study did not show any correlation between distribution of the diverse potential urovirulence factors and isolate source. No significant correlation was observed between infectivity and the origin of the isolates, since they all similarly colonized the urinary tract of the challenged mice. Finally, all isolates showed unique ERIC-PCR patterns, indicating that the isolates were genetically diverse. The results obtained in this study suggest that the source of P. mirabilis strains cannot be correlated with pathogenic attributes, and that the distribution of virulence factors between isolates of different origins may correspond to the opportunistic nature of the organism.
-
-
- Physiology
-
-
-
Osmotic stress in Synechocystis sp. PCC 6803: low tolerance towards nonionic osmotic stress results from lacking activation of glucosylglycerol accumulation
More LessIn order to compare the molecular principles of the acclimatization of bacterial cells to salt and nonionic osmotic stress, the moderately halotolerant cyanobacterium Synechocystis sp. PCC 6803 was challenged by salt (NaCl), and the osmolytes sorbitol and maltose. The physiological response towards each of the three compounds was found to be different. After salt addition, the cell volume remained unchanged, and the accumulation of the osmoprotective compound glucosylglycerol (GG) was observed after activation of the key enzyme GgpS at the biochemical and gene (ggpS) expression level. Sorbitol addition had only minor effects on the cell volume. In spite of the fact that the ggpS expression was increased, the GgpS enzyme was not activated, resulting in the absence of GG accumulation. In contrast the cells accumulated sorbitol, which served as a compatible solute and assured a certain osmotic resistance. In comparison to NaCl and sorbitol, the addition of maltose caused a strong decrease in cell volume indicating water efflux. However, no osmolyte accumulation was observed, resulting in an osmosensitive phenotype. Consequently, a successful response of Synechocystis cells to an osmotic challenge is indicative of the de novo synthesis of GG upon salt-dependent activation of the GgpS enzyme or the uptake of external solutes.
-
-
-
-
Candida albicans SNO1 and SNZ1 expressed in stationary-phase planktonic yeast cells and base of biofilm
More LessThe Candida albicans homologues of the most studied Saccharomyces cerevisiae stationary-phase genes, SNO1 and SNZ1, were used to test the hypothesis that, within a biofilm, some cells reach stationary phase within continuously fed, as well as static, C. albicans biofilms grown on dental acrylic. The authors first studied the expression patterns of these two genes in planktonic growth conditions. Using real-time RT-PCR (RT-RTPCR), increased peak expression of both SNZ1 and SNO1 was observed at 5 and 6 days, respectively, in C. albicans grown in suspension culture. SNZ1–yellow fluorescent protein (YFP) and SNO1–YFP were constructed to study expression at the cellular level and protein localization in C. albicans. Snz1p–YFP and Sno1p–YFP localized to the cytoplasm with maximum expression (>90 %) at 5 and 6 days, respectively, in planktonic conditions. When yeast growth was reinitiated, loss of fluorescence began immediately. Germ tubes and hyphae were non-fluorescent. Pseudohyphae began appearing at 9 days in planktonic yeast culture and expressed each protein by 11 days; however, the cells budding from pseudohyphae were not fluorescent. Biofilm was formed in vitro under either static or continuously fed conditions. Increased expression of the two genes was shown by RT-RTPCR, beginning by day 3 and increasing through to day 15 (continuously fed biofilm). Only the bottommost layer of acrylic-adhered cells in the biofilm showed 25 and 40 % fluorescence at 6 and 15 days, respectively. These observations suggest that only a few cells in C. albicans biofilms express genes associated with the planktonic stationary phase and that these are found at the bottom of the biofilm adhered to the surface.
-
-
-
Effect of AmtB homologues on the post-translational regulation of nitrogenase activity in response to ammonium and energy signals in Rhodospirillum rubrum
More LessThe AmtB protein transports uncharged NH3 into the cell, but it also interacts with the nitrogen regulatory protein PII, which in turn regulates a variety of proteins involved in nitrogen fixation and utilization. Three PII homologues, GlnB, GlnK and GlnJ, have been identified in the photosynthetic bacterium Rhodospirillum rubrum, and they have roles in at least four overlapping and distinct functions, one of which is the post-translational regulation of nitrogenase activity. In R. rubrum, nitrogenase activity is tightly regulated in response to
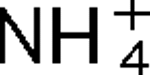 addition or energy depletion (shift to darkness), and this regulation is catalysed by the post-translational regulatory system encoded by draTG. Two amtB homologues, amtB
1 and amtB
2, have been identified in R. rubrum, and they are linked with glnJ and glnK, respectively. Mutants lacking AmtB1 are defective in their response to both
addition or energy depletion (shift to darkness), and this regulation is catalysed by the post-translational regulatory system encoded by draTG. Two amtB homologues, amtB
1 and amtB
2, have been identified in R. rubrum, and they are linked with glnJ and glnK, respectively. Mutants lacking AmtB1 are defective in their response to both  addition and darkness, while mutants lacking AmtB2 show little effect on the regulation of nitrogenase activity. These responses to darkness and
addition and darkness, while mutants lacking AmtB2 show little effect on the regulation of nitrogenase activity. These responses to darkness and 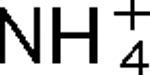 appear to involve different signal transduction pathways, and the poor response to darkness does not seem to be an indirect result of perturbation of internal pools of nitrogen. It is also shown that AmtB1 is necessary to sequester detectable amounts GlnJ to the cell membrane. These results suggest that some element of the AmtB1-PII regulatory system senses energy deprivation and a consistent model for the integration of nitrogen, carbon and energy signals by PII is proposed. Other results demonstrate a degree of specificity in interaction of AmtB1 with the different PII homologues in R. rubrum. Such interaction specificity might be important in explaining the way in which PII proteins regulate processes involved in nitrogen acquisition and utilization.
appear to involve different signal transduction pathways, and the poor response to darkness does not seem to be an indirect result of perturbation of internal pools of nitrogen. It is also shown that AmtB1 is necessary to sequester detectable amounts GlnJ to the cell membrane. These results suggest that some element of the AmtB1-PII regulatory system senses energy deprivation and a consistent model for the integration of nitrogen, carbon and energy signals by PII is proposed. Other results demonstrate a degree of specificity in interaction of AmtB1 with the different PII homologues in R. rubrum. Such interaction specificity might be important in explaining the way in which PII proteins regulate processes involved in nitrogen acquisition and utilization.
-
-
-
Global gene expression in Escherichia coli K-12 during short-term and long-term adaptation to glucose-limited continuous culture conditions
More LessMicroarray technology was used to study the cellular events that take place at the transcription level during short-term (physiological) and long-term (genetic) adaptation of the faecal indicator bacterium Escherichia coli K-12 to slow growth under limited nutrient supply. Short-term and long-term adaptation were assessed by comparing the mRNA levels isolated after 40 or 500 h of glucose-limited continuous culture at a dilution rate of 0.3 h−1 with those from batch culture with glucose excess. A large number of genes encoding periplasmic binding proteins were upregulated, indicating that the cells are prepared for high-affinity uptake of all types of carbon sources during glucose-limited growth in continuous culture. All the genes belonging to the maltose (mal/lamB) and galactose (mgl/gal) operons were upregulated. A similar transcription pattern was observed for long-term cultures except that the expression factors were lower than in the short-term adaptation. The patterns of upregulation were confirmed by real-time RT-PCR. A switch from a fully operational citric acid cycle to the PEP-glyoxylate cycle was clearly observed in cells grown in glucose-limited continuous culture when compared to batch-grown cells and this was confirmed by transcriptome analysis. This transcriptome analysis confirms and extends the observations from previous proteome and catabolome studies in the authors' laboratory.
-
-
-
Functional identification of ygiP as a positive regulator of the ttdA-ttdB-ygjE operon
More LessFunctional characterization of unknown genes is currently a major task in biology. The search for gene function involves a combination of various in silico, in vitro and in vivo approaches. Available knowledge from the study of more than 21 LysR-type regulators in Escherichia coli has facilitated the classification of new members of the family. From sequence similarities and its location on the E. coli chromosome, it is suggested that ygiP encodes a lysR regulator controlling the expression of a neighbouring operon; this operon encodes the two subunits of tartrate dehydratase (TtdA, TtdB) and YgiE, an integral inner-membrane protein possibly involved in tartrate uptake. Expression of tartrate dehydratase, which converts tartrate to oxaloacetate, is required for anaerobic growth on glycerol as carbon source in the presence of tartrate. Here, it has been demonstrated that disruption of ygiP, ttdA or ygjE abolishes tartrate-dependent anaerobic growth on glycerol. It has also been shown that tartrate-dependent induction of the ttdA-ttdB-ygjE operon requires a functional YgiP.
-
- Plant-Microbe Interactions
-
-
-
Antimicrobial activity of potato aspartic proteases (StAPs) involves membrane permeabilization
More LessSolanum tuberosum aspartic proteases (StAPs) with antimicrobial activity are induced after abiotic and biotic stress. In this study the ability of StAPs to produce a direct antimicrobial effect was investigated. Viability assays demonstrated that StAPs are able to kill spores of Fusarium solani and Phytophthora infestans in a dose-dependent manner. Localization experiments with FITC-labelled StAPs proved that the proteins interact directly with the surface of spores and hyphae of F. solani and P. infestans. Moreover, incubation of spores and hyphae with StAPs resulted in membrane permeabilization, as shown by the uptake of the fluorescent dye SYTOX Green. It is concluded that the antimicrobial effect of StAPs against F. solani and P. infestans is caused by a direct interaction with the microbial surfaces followed by membrane permeabilization.
-
-
-
-
Isolation of salt-sensitive mutants of Sinorhizobium meliloti strain Rm1021
More LessThe determinants necessary for adaptation to high NaCl concentrations and competition for nodule occupancy in Sinorhizobium meliloti were investigated genetically. Mutations in fabG as well as smc02909 (transmembrane transglycosylase), trigger factor (tig) and smc00717 (probably ftsE) gave rise to strains that were unable to tolerate high salt and were uncompetitive for nodule occupancy relative to the wild-type. Moreover exoF1, exoA and pgm determinants were determined to be necessary for strain Rm1021 to survive high NaCl and/or MgCl2 concentrations. The introduction of an expR + allele was capable of suppressing the Mg2+ sensitivity associated with the exoF1, but not the exoA, mutation in a manner independent of exopolysaccharide II (EPS II)-associated mucoidy. The results also show that the EPS II-associated mucoid phenotype was affected by either Mg2+or K+, but not by Li+, Ca2+, or high osmolarity.
-
-
-
Characterization of genes involved in erythritol catabolism in Rhizobium leguminosarum bv. viciae
More LessA genetic locus encoding erythritol uptake and catabolism genes was identified in Rhizobium leguminosarum bv. viciae, and shown to be plasmid encoded in a wide range of R. leguminosarum strains. A Tn5-B22 mutant (19B-3) unable to grow on erythritol was isolated from a mutant library of R. leguminosarum strain VF39SM. The mutated gene eryF was cloned and partially sequenced, and determined to have a high homology to permease genes of ABC transporters. A cosmid complementing the mutation (pCos42) was identified and was shown to carry all the genes necessary to restore the ability to grow on erythritol to a VF39SM strain cured of pRleVF39f. In the genomic DNA sequence of strain 3841, the gene linked to the mutation in 19B-3 is flanked by a cluster of genes with high homology to the known erythritol catabolic genes from Brucella spp. Through mutagenesis studies, three distinct operons on pCos42 that are required for growth on erythritol were identified: an ABC-transporter operon (eryEFG), a catabolic operon (eryABCD) and an operon (deoR-tpiA2-rpiB) that encodes a gene with significant homology to triosephosphate isomerase (tpiA2). These genes all share high sequence identity to genes in the erythritol catabolism region of Brucella spp., and clustalw alignments suggest that horizontal transfer of the erythritol locus may have occurred between R. leguminosarum and Brucella. Transcription of the eryABCD operon is repressed by EryD and is induced by the presence of erythritol. Mutant 19B-3 was impaired in its ability to compete against wild-type for nodulation of pea plants but was still capable of forming nitrogen-fixing nodules.
-
Volumes and issues
-
Volume 170 (2024)
-
Volume 169 (2023)
-
Volume 168 (2022)
-
Volume 167 (2021)
-
Volume 166 (2020)
-
Volume 165 (2019)
-
Volume 164 (2018)
-
Volume 163 (2017)
-
Volume 162 (2016)
-
Volume 161 (2015)
-
Volume 160 (2014)
-
Volume 159 (2013)
-
Volume 158 (2012)
-
Volume 157 (2011)
-
Volume 156 (2010)
-
Volume 155 (2009)
-
Volume 154 (2008)
-
Volume 153 (2007)
-
Volume 152 (2006)
-
Volume 151 (2005)
-
Volume 150 (2004)
-
Volume 149 (2003)
-
Volume 148 (2002)
-
Volume 147 (2001)
-
Volume 146 (2000)
-
Volume 145 (1999)
-
Volume 144 (1998)
-
Volume 143 (1997)
-
Volume 142 (1996)
-
Volume 141 (1995)
-
Volume 140 (1994)
-
Volume 139 (1993)
-
Volume 138 (1992)
-
Volume 137 (1991)
-
Volume 136 (1990)
-
Volume 135 (1989)
-
Volume 134 (1988)
-
Volume 133 (1987)
-
Volume 132 (1986)
-
Volume 131 (1985)
-
Volume 130 (1984)
-
Volume 129 (1983)
-
Volume 128 (1982)
-
Volume 127 (1981)
-
Volume 126 (1981)
-
Volume 125 (1981)
-
Volume 124 (1981)
-
Volume 123 (1981)
-
Volume 122 (1981)
-
Volume 121 (1980)
-
Volume 120 (1980)
-
Volume 119 (1980)
-
Volume 118 (1980)
-
Volume 117 (1980)
-
Volume 116 (1980)
-
Volume 115 (1979)
-
Volume 114 (1979)
-
Volume 113 (1979)
-
Volume 112 (1979)
-
Volume 111 (1979)
-
Volume 110 (1979)
-
Volume 109 (1978)
-
Volume 108 (1978)
-
Volume 107 (1978)
-
Volume 106 (1978)
-
Volume 105 (1978)
-
Volume 104 (1978)
-
Volume 103 (1977)
-
Volume 102 (1977)
-
Volume 101 (1977)
-
Volume 100 (1977)
-
Volume 99 (1977)
-
Volume 98 (1977)
-
Volume 97 (1976)
-
Volume 96 (1976)
-
Volume 95 (1976)
-
Volume 94 (1976)
-
Volume 93 (1976)
-
Volume 92 (1976)
-
Volume 91 (1975)
-
Volume 90 (1975)
-
Volume 89 (1975)
-
Volume 88 (1975)
-
Volume 87 (1975)
-
Volume 86 (1975)
-
Volume 85 (1974)
-
Volume 84 (1974)
-
Volume 83 (1974)
-
Volume 82 (1974)
-
Volume 81 (1974)
-
Volume 80 (1974)
-
Volume 79 (1973)
-
Volume 78 (1973)
-
Volume 77 (1973)
-
Volume 76 (1973)
-
Volume 75 (1973)
-
Volume 74 (1973)
-
Volume 73 (1972)
-
Volume 72 (1972)
-
Volume 71 (1972)
-
Volume 70 (1972)
-
Volume 69 (1971)
-
Volume 68 (1971)
-
Volume 67 (1971)
-
Volume 66 (1971)
-
Volume 65 (1971)
-
Volume 64 (1970)
-
Volume 63 (1970)
-
Volume 62 (1970)
-
Volume 61 (1970)
-
Volume 60 (1970)
-
Volume 59 (1969)
-
Volume 58 (1969)
-
Volume 57 (1969)
-
Volume 56 (1969)
-
Volume 55 (1969)
-
Volume 54 (1968)
-
Volume 53 (1968)
-
Volume 52 (1968)
-
Volume 51 (1968)
-
Volume 50 (1968)
-
Volume 49 (1967)
-
Volume 48 (1967)
-
Volume 47 (1967)
-
Volume 46 (1967)
-
Volume 45 (1966)
-
Volume 44 (1966)
-
Volume 43 (1966)
-
Volume 42 (1966)
-
Volume 41 (1965)
-
Volume 40 (1965)
-
Volume 39 (1965)
-
Volume 38 (1965)
-
Volume 37 (1964)
-
Volume 36 (1964)
-
Volume 35 (1964)
-
Volume 34 (1964)
-
Volume 33 (1963)
-
Volume 32 (1963)
-
Volume 31 (1963)
-
Volume 30 (1963)
-
Volume 29 (1962)
-
Volume 28 (1962)
-
Volume 27 (1962)
-
Volume 26 (1961)
-
Volume 25 (1961)
-
Volume 24 (1961)
-
Volume 23 (1960)
-
Volume 22 (1960)
-
Volume 21 (1959)
-
Volume 20 (1959)
-
Volume 19 (1958)
-
Volume 18 (1958)
-
Volume 17 (1957)
-
Volume 16 (1957)
-
Volume 15 (1956)
-
Volume 14 (1956)
-
Volume 13 (1955)
-
Volume 12 (1955)
-
Volume 11 (1954)
-
Volume 10 (1954)
-
Volume 9 (1953)
-
Volume 8 (1953)
-
Volume 7 (1952)
-
Volume 6 (1952)
-
Volume 5 (1951)
-
Volume 4 (1950)
-
Volume 3 (1949)
-
Volume 2 (1948)
-
Volume 1 (1947)
Most Read This Month
